シダ植物ヘビノネゴザにおける重金属耐性と蓄積に関する研究

シダ植物のヘビノネゴザは、日本各地の金属鉱山や重金属汚染地にしばしば群生しており、重金属毒性に高い耐性を持っているだけでなく植物体にカドミウム、鉛、銅、亜鉛などの重金属を高濃度に蓄積する性質がある。我々はこのようなヘビノネゴザの性質に興味を持ち、以下のような研究を行なっている。
配偶体における重金属耐性と重金属集積性
シダ植物は配偶体世代(有性世代)と胞子体世代(無性世代)の植物体が独立して生活している。我々は胞子発芽を指標にしてヘビノネゴザ配偶体にも重金属に対する耐性が存在し、仮根に鉛を多量に蓄積することを明らかにした。また、ヘビノネゴザ配偶体にはプロアントシアニジン(縮合型タンニン)が高濃度に存在することを明らかにし、プロアントシアニジンが鉛耐性に関与することを示唆した1。そこで、プロアントシアニジンの含有量が低下した配偶体(Aypad1)を作製し、鉛とカドミウムに対する耐性と蓄積について調べた。しかし、Aypad1配偶体のこれら重金属に対する耐性と蓄積能は野生型(WT)配偶体のものと有意差は見られず2、現在検討を続けている。
1Kamachi H, Komori I,Tamura H, Sawa Y, Karahara I, Honma Y, Wada N, Kawabata T, Matsuda K, Ikeno S, Noguchi M and Inoue H (2005) Lead tolerance and accumulation in the gametophytes of the fern Athyrium yokoscense. Journal of Plant Research 118:137-145.
2Kamachi H, Morishita K, Hatta M, Okamoto A, Fujii K, Imai N, Sakatoku A, Ohta T, Aoki M, Hiyama S (2021) Lead and cadmium tolerance and accumulation of proanthocyanidin-deficient mutants of the fern Athyrium yokoscense. International Journal of Plant Biology 12:9330
バリウム (Ba) の集積性と耐性
ヘビノネゴザは、カドミウム、鉛、銅、亜鉛以外の重金属は蓄積しないのだろうか?野外からヘビノネゴザを採取し、ICP発光分光分析法による多元素分析を行ったところ、葉身にバリウムを蓄積 (Max. 1,000 ppm) する性質を持つことが、新たにわかった。植物にって、高濃度のバリウムはカリウムの取り込みを阻害し植物の成長を抑制するが、ヘビノネゴザでは、バリウム毒性に対しても耐性を示すことが配偶体の培養実験から明らかとなった。
ヘビノネゴザは、他の植物と比較して、セシウム (Cs) を取り込みやすいことが報告されている。セシウム、バリウム、カリウムのイオン半径はよく似ていることから、ヘビノネゴザでは、カリウムチャネルが他の植物に比べて、バリウムイオンやセシウムイオンを通しやすい性質を持っているのかもしれない。
Kamachi H, Kitamura N, Sakatoku A, Tanaka D and Nakamura S (2015) Barium accumulation in the metalliferous fern Athyrium yokoscense. Theoretical and Experimental Plant Physiology 27(2):99-107.
リチャードミズワラビ配偶体の重力センシング
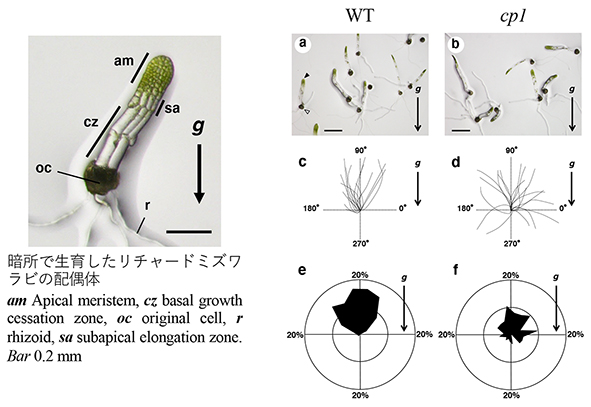
植物は重力方向を感知することができ、茎は重力と反対方向に、根は重力方向に成長する。重力方向を感知する細胞は平衡細胞と呼ばれ、デンプンの詰まった細胞小器官(アミロプラスト)が重力方向に沈降することにより、植物は重力方向を感知していると考えられている。これをデンプン平衡石説と呼ぶ。
シダ植物リチャードミズワラビの配偶体は、暗所で負の重力屈性を示したが、細胞内には平衡石と思われる様な沈降性の細胞小器官や物質は観察されず、非沈降性の平衡石が存在している可能性が考えられた1。葉緑体が細胞膜からはずれて凝集してしまう変異体(cp1)を用いて、葉緑体が重力感受に関係するかどうかを調べたところ、cp1 変異体の光屈性は正常であったにもかかわらず、重力屈性は大きく抑制されていた2。この結果は、リチャードミズワラビ配偶体では葉緑体が重力感受のための平衡石として機能しており、葉緑体が細胞膜へ固定されていることが重力感受に必要であることを示唆している。
1Kamachi H and Noguchi M (2012) Negative gravitropism in dark-grown gametophytes of the fern Ceratopteris richardii. American Fern Journal 102:147-153.
2Kamachi H, Tamaoki D, Karahara I (2017) Plasma membrane-anchored chloroplasts are necessary for the gravisensing system of Ceratopteris richardii prothalli. Journal of Plant Research130:397-405.
リチャードミズワラビ配偶体の性決定における光の影響

シダ植物では有性世代(配偶体世代)と無性世代(胞子体世代)の植物体がそれぞれ独立して生活しており、有性世代の配偶体には雌雄が存在する。シダ配偶体が確実に胞子体を作る生存戦略として、性比を性フェロモン(アンセリジオーゲン)によって決定するやり方が知られている。つまり早く発芽した胞子が雌あるいは雌雄同体の配偶体へと成長しアンセリジオーゲンを周囲に分泌するようになるため、遅れて発芽した胞子は雄へと成長する。したがって、雌性配偶体の周囲には多数の雄性配偶体が形成され受精がより確実に行われる。なお、雌性配偶体はアンセリジオーゲンの影響は受けず、また雄性配偶体でもアンセリジオーゲンがなくなるとやがて雌性配偶体へと成長する。
本研究では、リチャードミズワラビの性決定機構に光環境がどのように影響するのかを調べ以下の結論を得た。(1) 青色光はアンセリジオーゲンの存在下において配偶体の雄性化を若干促進する作用をもつ。(2) 赤色光には配偶体を雌性化する作用があり、この作用は(1)の青色光効果よりも優位に作用する。(3) (2)の赤色光効果はフィトクロムによるものである。
植物は、赤色光の割合の高い光環境を光合成に有利な光環境であると判断するため、そのような光環境下では雌性配偶体がより多く形成され、その結果胞子体もより多く形成されるものと思われる。このように、シダ配偶体の性決定には光環境も影響していることがわかった。
Kamachi H, Iwasawa O, Hickok LG, Nakayama M, Noguchi M and Inoue H (2007) The effects of light on sex determination in gametophytes of the fern Ceratopteris richardii. Journal of Plant Research 120:629–634.
リチャードミズワラビ配偶体の多様なフィトクロム反応
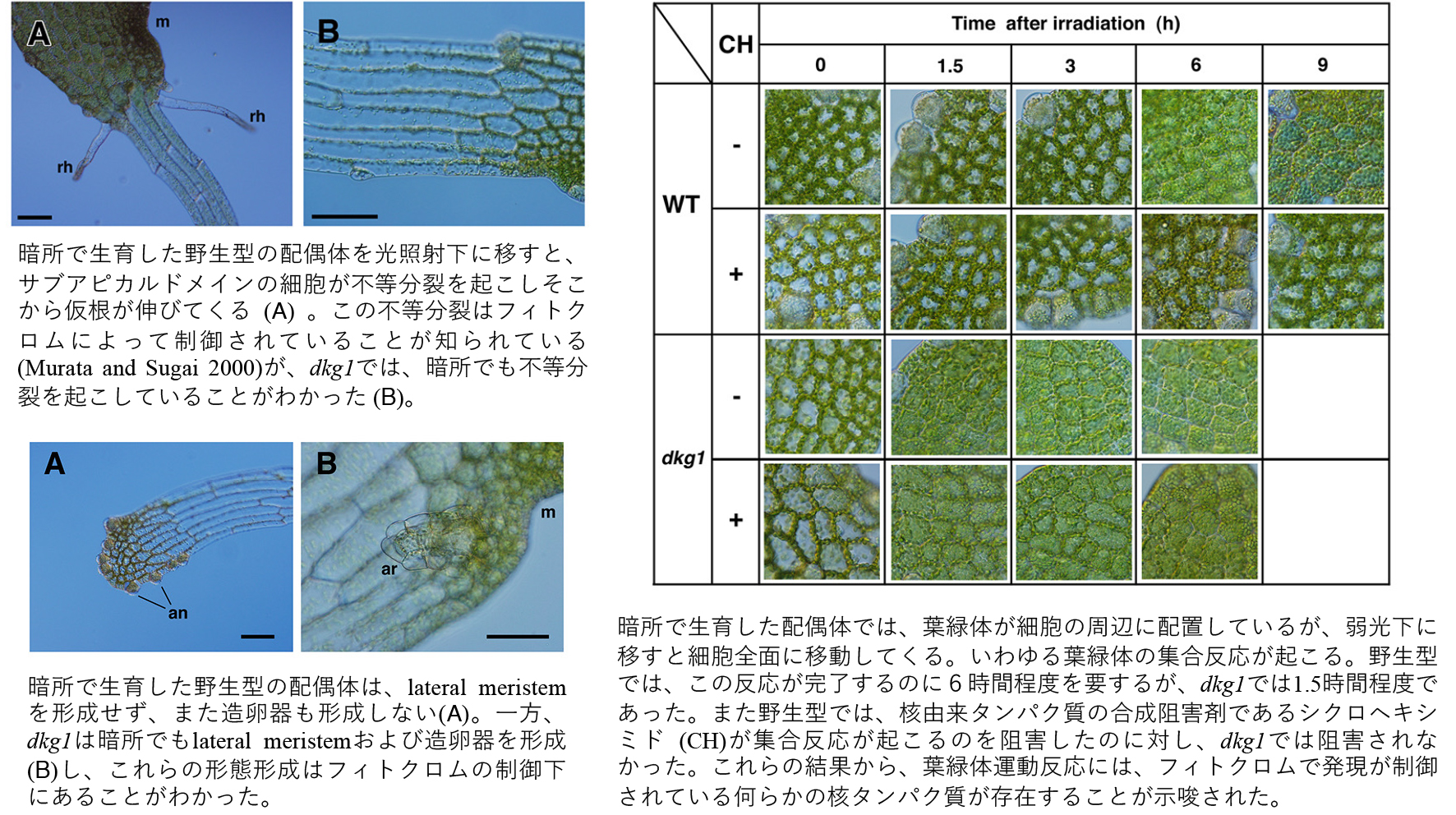
シダ胞子の光発芽は典型的なフィトクロム反応を示す。本研究では、リチャードミズワラビの暗所胞子発芽突然変異体 (dkg1) の配偶体の成長過程を詳細に観察し、配偶体の様々な生理応答がフィトクロム制御下にあることを報告した。
Kamachi H, Matsunaga E, Noguchi M and Inoue H (2004) Novel mutant phenotypes of a dark-germinating mutant dkg1 in the fern Ceratopteris richardii. Journal of Plant Research 117:163-170.
ゼンマイ胞子の光発芽過程におけるカルシウム情報伝達
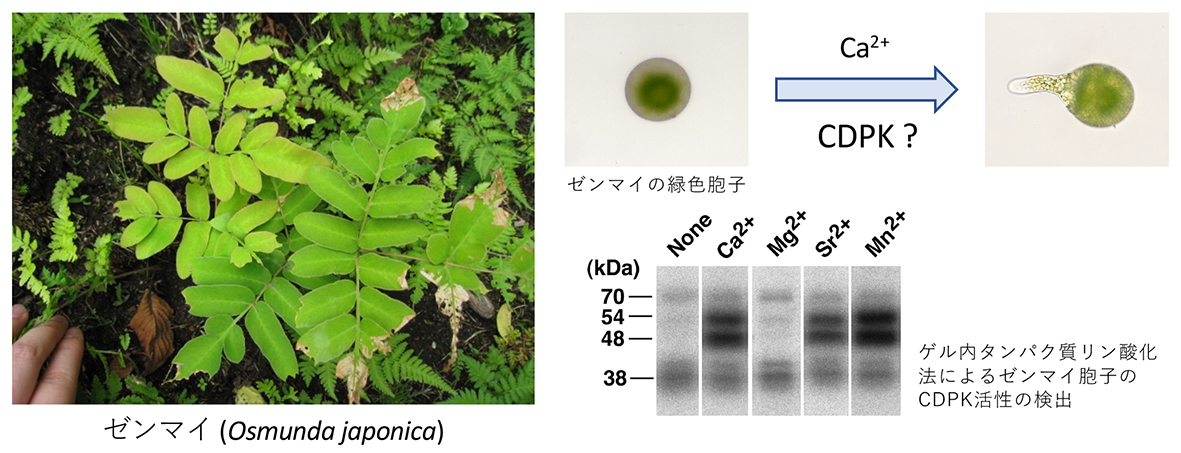
シダの胞子発芽にはカルシウムイオンが必須である。本研究では、分子量が54と48 kDaのカルシウムイオン依存性プロテインキナーゼ(CDPK)が、ゼンマイの胞子発芽に関与する可能性を示した。
Kamachi H, Noguchi M and Inoue H (2004) Possible involvement of Ca2+-dependent protein kinases in spore germination of the fern Osmunda japonica, Journal of Plant Biology, 47(1):27-32.